
lobe ode ear node


Over this last while I’ve written a number of essays who refer to human ear anatomy, so here I am trying to consolidate some notes I’ve made over years, perhaps for no other reason than exfoliating an overly mechanistic layer of my sensibility.
This was my hope whilst writing Azimuth, but information’s abundance seemed better suited to a poetics. Here I try to focus on who we could call a practical nature, at best a brief introduction, though there will surely be occasion to drift, and certainly to return, as ear is both a good place to sleep and to leave. I can’t imagine many reading this through, but I hope it can provide a frame of reference when needed.
Perhaps some will note absence, or scarcity, of information regarding such creatures as temporal bone and auditory bulla. I leave them out purely because if I include them I will then need to include someone else and so on. I also haven’t included a list of references because said list would be a giant. However, please feel free to get in touch if there’s anything specific you’d like to talk about, hopefully I can point in a direction.
It’s difficult to avoid generalising, though when I do, it is more for ease and use than anybody else, as during such moments I’m not talking about myself (my ear is anyone but general), so in essence I could be talking about anyone and any one.
Likewise, devising any semblance of coherent order is essentially an exercise in superficiality and hodology/pathology. We could go any number of ways, especially in mind of quavering ultrastructures of labyrinths, liable to flip at any moment, like Earth’s magnetic fields.
This cornucopia is brought to light in relation to someone resembling aural diversity inasmuch as these notes will not only hear themselves differently, but will change their meaning in accordance the reader. This is closely akin to Felix Guatarri’s notion of metaphor as metamorphosis, a displacement who produces a prospective reconfiguration of sense produced by symptom between word and person, word between person and symptom, and so on. Like birds who come and tap windows.
I feel like I’ve written with human ears (from hereon, ‘ear’ will denote human ear) more times than I can recall, yet every time I do, I encounter someone I’d previously missed, or I encounter new research. As such, without wishing to repeat myself, please consider this a process.
I’ve tried many times to open ear into a much wider space of ideas, a space not predicated on protein and voltage’s vernacular. This is why I keep coming back, I sleep in ear because there’s always more to hear (to remember I’ve already forgotten) about their micro and macro molecular natures and substructures, molecular warp and woof.
By ‘opening’ ear, I’m referring to their many lives (not only human)... subtle, spiritual, electric and magnetic, their noise and gravitation, sound, heat and light… innumerable chemical signals, their energy! Amidst this sweating feedback loop who drops their own boundaries we need also include affect and emotion, who else… all sorts of pullulating myths as they metamorphose in such densely spectral labyrinths, guided by slipping in and out of morphic wholeness, where one being is always on their way to becoming another. The faster such forces follow one another the more coherent a continuum they form as they in turn move further apart.
Ear is immanent and innumerable, one might be abandoned, one might catch fire, another might be carried way by the wind, another might vent like a spleen. Perhaps it’s more prudent to speak about phases and maturations, rather than endings and fructifications.
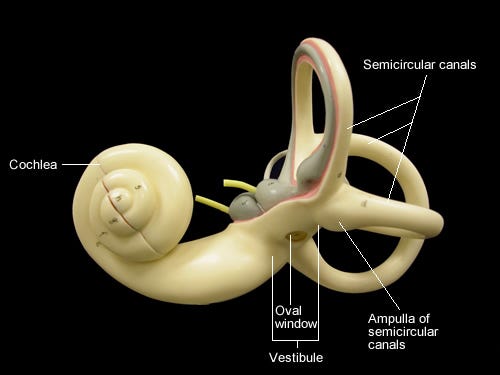
Auricle, (’outer’ ear) is unique in their morphology, inasmuch as they act as mirror for who they makes sound appear to be. Sound created ear in order to hear themselves, that old adage. Creature populate their surface, helix, tragus, concha, and grow their venation.
Auricle twists and folds in such a way that they enhance sound within seams typical of human voices (much like tectorial membrane). Their shape is such that they can amplify vibrations up to 100 times. Vibrations from ‘front’ and ‘sides’ are enhanced in such a way, those circling from back are reduced, creating small differences in volume and a tendency for body to participate in their own asymmetrical spectral reflection.
Auricle means ‘ear shaped’, curious to think that outer ear is called an auricle because outer ear looks like an outer ear.
Auricle can also be referred to as pinna, Latin for wing. Pinna appears in botany as primary segment of compound leaf, a genus of bivalve molluscs known as pen-shells, and in zoology as a feather or fin. Auricle emerges from auditory, who themselves relate to medieval rituals of confession. Language and experience can reverse, creating a kind of wild logos, a transmission who is also a reception.
We listen to oracle we confess to auricle, and so auditory is ancestor of auricularis, relating to any three muscles, triangle attaching cartilage of external ear to skull. Auricular can mean anyone relating to ear, or hearing. Tracing these vagaries and disturbed certainties, evolution of a word is akin to a sound wave mapping and expanding their own self-similarity.
Ear canal ( auditory canal / external auditory meatus) consists primarily of bone, fibre, and cartilage. They are self cleaning, if we consider that cerumen is part of their anatomy (and that they are part of cerumen’s) and they are often around 2.5cm long, 0.7 in diameter. There is usually a slight curve in their tube, extending from pinna to arrive and depart at tympanic membrane.
Tympanic membrane (tympanum / ear drum) is a thin and oscillating transparent partition of nonglandular skin who separates and links auditory canal from depths of middle ear cavity. As such, they both receive and transmit environmental vibration, from where we happen to be in relation with excitations of ossicles.
They are stretched obliquely (slanting) from above to below (such attempts at orientation are relative), from outside to inside, and from back to front. Therefore they are not perpendicular to canal’s axis.
Tympanic membrane’s obliqueness increases surface impression and vibration’s capacity (ossicles amplify by a factor of three when sound comes into contact with liquid, like inner ear’s atmosphere, a great deal of whom is dispersed, so ossicles and tympanic membrane entangle impedance between air and fluid, and ossicles act in a similar fashion, contributing coupling required for vibration’s transmission about inner labyrinth’s floating world). Indeed, hearing’s precision is in direct proportion to tympanum’s obliqueness.
Membrane has three layers, outer - part of skin of the ear canal, inner - part of mucous membrane lining middle ear, and in between is a layer of radial and circular fibres who stiffen and tense proportions of disc. As such, they are vulnerable and sensitive to pain.
Membrane has been subject to predictable designations of proximity, apparently they measure difference in relation to whomsoever is ‘proper’ to oneself and obversely ‘belongs’ in ‘other’s’ realm. I would rather concentrate on their vocabulary, as disc is home to such terms as umbo, as most depressed point, who, put simply, corresponds to malleus (ossicle) and cone of light, who acts as a guide upon closer audiological intimacy.
Anatomically speaking, Eustachian tube is, in their own way, responsible for aforementioned proximity. They correspond with and connects middle ear to back of throat along whom we might call increments. Enabling air pressure (and draining fluid) to equalise on both sides of tympanic membrane’s film.
Middle ear’s air is not in ‘direct’ contact with atmosphere outside body, and so Eustachian tube can link many mediums via creation of pressure differentials. When we chew, yawn, and swallow, tube opens, otherwise they tend to remain closed. In other words, when we yawn we’re hearing tube open.
In 1546 Giovanni Filippo Ingrassia, professor of anatomy at Naples University (a then burgeoning city with roughly nine times the population of Marseille, marketplaces brimming with taffetas, silken knots and cockades, silken air promising freedom, but not necessarily a full stomach, identified a third small bone in middle ear, an ossified ring he named stapes. He writes:
By chance, while showing students the two small bones of the middle ear, the malleus and the incus, I noticed that a third bone had fallen on the dissecting table, and for its similarity to a bracket or to the Greek letter Delta, I thought to call it ‘stapes’ or ‘deltoid.’
Ingrassia was called a scholar of human’s body, he lectured on medicinal theory and practise, on Avicenna and al-Razi. Possessing a rare osteo-sensibility, he gained even greater renown upon declaring that Galen, famous philosopher-surgeon of Ancient Rome, had often described monkey bones when writing about those of humans.
Ingrassia intuited symbolic wildness and under-textures of inner ear’s oval and round windows, hypothesised on vibratory capacity of teeth, opined extra-sensorial bone conduction.
Ossicles (malleus/incus/stapes) are body’s smallest bones, they evolved from lower jawbone of amniotes and transduce vibration from one medium to another. Suspended by ligaments, they form a path from ear to ear. When arranged together, ossicles look like a rhizome of ginger and move like a moorhen washing in muddy water.
Malleus (hammer) lifts from tympanic membrane within the middle ear’s cavern. They’re attached to incus (anvil). Malleus has a neck and evolved from vertebrates who laid their eggs on land. Their handle is sometimes noted as a reddish-yellow streak. If we traverse bone as a whole, or part of ossicular line, we note that their extreme end, in mind of their infinitesimal nature, is a little white and rounded prominence.
Middle ear once had a function to support jaw. Malleus sprouted from jaw joint, they now have a long arm and a bony wing. Incus receives lateral vibration from malleus, which they then transmit to stapes (stirrup).
Ossicles are a distortion of many bodies, many environments, stapes wants to break a window who wants to be broken, yet they cannot, because they are body’s smallest and lightest bone.
Middle ear responses are influenced by inner ear’s ability to process frequencies being transmitted. Where ossicles are called a chain, they can also be called a line. For many therian mammals middle ear is encased in a bony structure called auditory bulla, a dark space, floor of middle ear’s cavity. This protects middle ear tissue–they are also a genus of hermaphrodite sea snails, and Spanish for noise.
Oval window is a reniform (kidney-shaped) membrane, a covered aperture through whom vibrations travel from ossicular pivot of middle ear, they are yet another intersection in an ear seemingly made up of no one else.
Any one part of cochlea is difficult to articulate without then needing to refer to others, and to others in turn. At this point, detail’s depth is essentially limitless, and orientation becomes increasingly difficult. When I read about inner ear anatomy and auditory pathways I a reading a science fiction novel.
Ear is a fractal organ who is part of fractal body. To look into labyrinths is a journey of imagination, to enter into an archaic revival, which is not to say, going back to someplace or somewhere, not a return to, but a return of. In cochlea’s spiral is cosmos.
Cochlea is a fluid filled spiral within a fluid filled spiral who never really looks like a spiral because their edges are always in motion. They span roughly two and a half revolutions in humans, this corresponds to a frequency spectrum, and is otherwise known as auditory (osseous) labyrinth (within whom we find membranous labyrinth). Settled on cochlea’s spiral are many names, and those names also have other names, I’m going to try and stick to just one set.
As air pressure fluctuations and all they contain pass through cochlea (initially known as vestibular duct), stapes causes oval window to compress, this in turn, at cochlea’s alternate end (tympanic duct), causes round window to move in another direction, this works both ways in compression (causing an increase in local cochlear pressure) and rarefaction (causing a decrease). This is to create a fluid pressure equilibrium.
This stimulation can happen also via direct vibration of cochlea from skull, a form of bone conduction hearing, which can be complementary (and not) to air conduction hearing. Both AC and BC stimulate basilar membrane of cochlea similarly. Here we need note that basilar membrane moves in accord with longitudinal wave patterns, so with compression and rarefaction, whilst tectorial membrane can move as transverse wave, who crests and troughs. This is of particular interest when we note that longitudinal wave forms are traditionally associated with sound, and transverse with electromagnetism, of light.
A recent experiment out of City university in Hong Kong has shown however, that sound can behave like a transverse wave. I bring this up because of alchemical possibility, by a shift in perspective, apparent opposites of someone may come into focus at any moment. Many science fiction writers, notably Alice Bradley Sheldon, otherwise known as James Tiptree Jr. pre-empted such findings by referring to such phenomena as light-sound, most notably in her book, Up the Walls of the World, published in 1978.
Just to recap. Waves of metamorphosing pressure come through oval window into vestibular duct, pass around spiral to apex of osseous labyrinth, through who is known as helicotrema and into tympanic duct, leading to round window. This is a continuous passage of fluid and pressure.
Both vestibular and tympanic ducts of osseous labyrinth are full of perilymph, who is essentially a conductive fluid. In between these ducts is another spiral, called cochlear duct, this gets a little complicated when we realise that apex of cochlear duct is at base of the osseous cochlea. Cochlear duct is full of endolymph, a unique conductive fluid rich in potassium.
What’s currently known as tonotopy (tonos - tone / topos - place) is a mapping of different frequencies along cochlea length. This was ‘discovered’ by George von Békésy mid 20th century. Tonotopy is thought to begin at cochlea, and then spreads throughout brain, though I feel they’ll be observed throughout body, and indeed, why stop there... This is part of a much wider field of auricular therapy and Meridians, which I hope, is to come.
When we consider inner ear’s tonotopic life we are helping to tune and be tuned by multi-dimensional auricular branches. High frequencies congregate amongst cochlea’s base, and lowest flow amongst apex, so again, this is opposite to what we’d logically deduce… and has to do, at least in part, with inverted relationships of three ducts.
I should at least briefly mention cochlear amplifier, who increases deflection of membranes in cochlear duct, but also augments resolution who we can detect between two different frequencies. So essentially, fluctuations who arrive as 1000 Hz will be amplified at one part of spiral, and those of 1005 Hz at another, this enables afferent and efferent motion of signals along auditory radiation pathways of brain.
Cochlea duct contains sensory region of cochlea, otherwise known as organ of Corti. Organ of Corti lies on basilar membrane at base of cochlear duct. Under organ of Corti is tympanic duct and above them is vestibular duct. The borders cochlea’s central bone, otherwise known as modiolus, and cochlea’s outside wall, who contains a tissue called stria vascularis, a very particular place pumping out endolymph. Here we recall that outside cochlear duct other two ducts (vestibular and tympanic) are bathed in perilymph, an extracellular fluid.
Stria vascularis also serves as a kind of battery, they charges endolymph, and this is called endocochlear potential, to around +80mv, which helps hair cells to respond to ‘environmental’ stimuli.
Basilar membrane separates tympanic and cochlear ducts (segregating endolymph and perilymph, though they are permeable to perilymph), undulating in response to pressure waves. They vary in width, stiffness, and mass along their length. Basilar membrane is widest and least stiff at cochlea’s apex, and subsequently stiff and narrow at their base. As we’ve said, high frequencies localise near base, and low near apex. Reissner’s membrane separates cochlear duct from vestibular duct, helping to transmit vibrations and nutrients between them.
Basilar membrane separates tympanic and cochlear ducts (segregating endolymph and perilymph, though they are permeable to perilymph), undulating in response to pressure waves. They vary in width, stiffness, and mass along their length. Basilar membrane is widest and least stiff at cochlea’s apex, and subsequently stiff and narrow at their base. As we’ve said, high frequencies localise near base, and low near apex. Reissner’s membrane separates cochlear duct from vestibular duct, helping to transmit vibrations and nutrients between them.
Cochlear duct is inside cochlea, inside cochlear duct is organ of Corti, inside said organ are hair-like cells, sensory cells releasing information onto spiral ganglion cells. In cochlear duct there are both inner and outer hair-like cells, inner hair (I’ll stop saying -like now) cells send messages to central nervous system, and outer hair cells in fact receive messages from brain. Outer hair cells are in essence, cochlear amplifier, who is a physical substrate enabling us to, well, as their name says, amplify sounds in cochlea.
All hair cells have cell bodies, there are also bunches of ‘hair-looking beings’, who are not hair at all, but whom we refer to as actin, formed as bundles, and outer hair cells (of whom there are approximately three times as many compared to inner hair cells) are embedded into tectorial membrane, who is made of molecules called tectorin. Inner bundles move within endolymph of organ of corti, and they of course move in a manner similar to oval and round windows.
If we try and map anatomy of these interconnected cells, there would cell body and stereocilia, joined by proteinaceous tiplinks, who join taller stereocilia to smaller (wherein tiplinks end), attached to met channel. When a bundle is moved we might say a door opens and ions can flow through said channel. Driving force along met channel goes from +80mv on endolymph’s side to around -50 mv on other side. This means hair cells have an elevated resting potential. Driving force is >100mv and that’s in stark contrast to a more typical force who is around 70mv. Once tip link’s lid comes off and channel is open, potassium, sodium and calcium all flow through cation channels, resulting in quick (about 20khz) responses.
Outer hair cells, embedded in tectorial membrane, amplify signals for inner hair cells, who are bathed in endolymph. They do this with a molecule called prestin, a molecular motor who changes their confirmation in response to voltage, shortening and lengthening cell lengths.
As waves of pressure pass through cochlea, outer hair cell bundles go back and forth in accord to compression and rarefaction of both windows, causing prestin molecules along hair cell to lengthen and shorten. This has a direct effect on inner hair cells, thereby creating stimulus. If there is no outer hair cell function, no prestin function, there is no hearing, as we understand them.
Békésy thought tonotopy was wave’s theory par excellence, thus explaining hearing, and yet now we have cochlear amplifier integrating into this particular vision, and on and on we shall go, continually breaking ear into pieces over and over and over again until eventually they become so small we lose ourselves in our own world.
Imagine all ear’s parts working in unison, from ripples and waves of conductive fluids to vibrations in bone to hair cells flowing back and forth, we are endless.
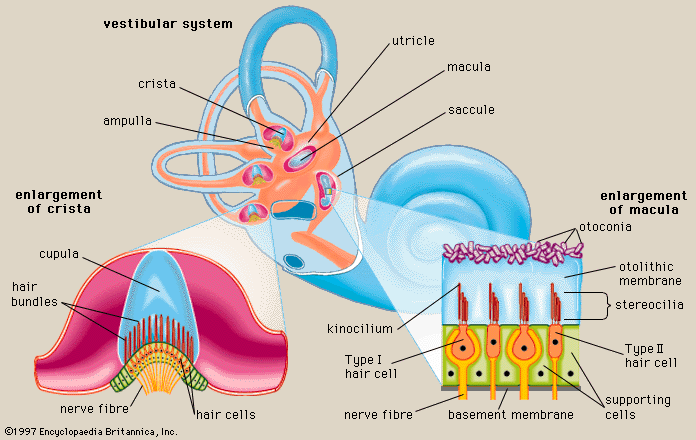
Human inner ear is a place of co-habitation, home to both cochlea and vestibulum. Whilst hearing and balance organs are linked, vibration should ‘never’ make fluid move in vestibulum.
A duct channels acoustic pressure changes away from balance organs so they can, for instance, send messages to brain concerning head’s position in relation to Earth’s gravitational fields.
Inner ear’s balance organs create vestibular labyrinth, who themselves consist of three semicircular canals and vestibule, who holds two otolith organs, utricle (closer to semicircular canals) and saccule (closer to cochlea). Angular acceleration is sensed by saccule, static head position in relation to Earth’s gravitational feels, and linear acceleration by utricule.
Semicircular canals are balance organs, and balance is a sense. Canals are tilt and gravity sensors who each correspond to a plane in which skull, and indeed body, can rotate. There are anterior (sometimes called superior), posterior and horizontal semicircular canals. They are all essentially concerned with rotational equilibrium.
Saccule and utricle are thin-walled membranous sacs and they contain neuroepithelium, a cluster of sensory and supporting cells aggregated in macula. Macula of utricle is in a horizontal position and in saccule, a vertical position.
Each cell contains stereocilia who project toward a sheet of gelatinous material, covering macular’s entire surface who in turn is covered by a mass of infinitesimal calcium carbonate crystals called otoconia, collectively constituting otoliths.
In the semicircular canals lives crista, who contains cupula. As stated, each otolith organs contain macula, full of otoconial crystals.
Otoconial layer, when loose, or unfixed, has a consistency of lightly packed sand, helping to project a steady stream of impulses around brain, even when head is at rest. When head moves, however slightly, layers shift in seams of minute displacement, deforming stereocilia, thus modulating neural messages.
Saccular crystals are fairly constant in size and uniform in shape, whereas utricular otoconia tend to be smaller and vary widely in shape, between rhombohedral and cylindrical. I would place otoconia in micromolecular structures of blades of straw, fractal distillates of time, continual turnover of calcium between otoconia and body’s calcium stores.
Crystals are not embedded in membrane’s gelatinous portion, only loosely bound by small amounts of matrix material, a surface of millions of small crystals and chemical interactions, so crystals can become dislodged, and drift.
A physiological mechanism exists in utricle for removal of dislodged otoconia. Crystals can come into contact with dark cells of utricular wall who appear capable of consuming displaced otoconia.
Frequencies of vestibular labyrinth are wildly different to those of auditory labyrinth. With human hearing, we’re of course dealing with a rough peak of 20 kHz. In vestibular system however, most stimulation is below 10 Hz, with head movements whilst walking being around 2-3 Hz.
As already stated, inside osseous labyrinth is membranous labyrinth, and between them is perilymph, a conduction fluid. Intimate anatomical proximity of balance and auditory organs in human body has led to a number of ways to guide acoustic pressure changes away from vestibular organs. Perilympthatic duct is one (where perilymphathic space is connected to subarachnoid space), differing resonant frequencies of vestibular and auditory labyrinths, another.
Otolith organs of inner ear have been called organs through whom gravity speaks. Within inner labyrinth, along a strange border between vestibular and auditory life, fluid and pervasive gravitational influence is most keenly felt.
Angular acceleration (how much someone spins, rotates, turns), is a talent of semicircular canals, in other words, rotational equilibrium. Linear acceleration (acceleration along a straight line) is a talent of otolith organs.
As already stated, at one end of each of canal there is crista, this is where hair cells are concentrated, oriented in one direction, embedded in a gelatinous veil, called cupula. There are three corresponding movements–yaw, pitch and roll–and these correspond not to a single canal, but register as mixture.
A yaw movement is a rotation in transverse plane, equivalent to horizontal canal, a side to side rotation of head, as though indicating ‘no’. As I practise this, fluid in membranous labyrinth (fixed to head) moves relative to head, creating a drag on cupula.
What this means is that if I move head in one direction, endolymph’s relative movement is in opposite direction. If I’m moving head right, endolymph in right ear excites hair cells, this is an equivalent of depolarisation. So each canal pairs, whomsoever happens in one, will counter their opposite.
If I move head a half pitch forward and a half roll right, that’ll make fluid in right anterior canal move away from utricle, so they’ll move opposite to head’s movement. Hair cells are oriented away from utricle, so whilst I move head one way, fluid moves their other way, and this excites hair cells, de-polarises them essentially. Subsequently hair cells left side of posterior canal will be inhibited, they’ll be hyper-polarised.
Virtually all movements are combinations, additions of at least two different vestibular stimuli, including saccule and utricle. When a person drinks a large amount of alcohol, specific gravity of blood goes down, cupula gets a blood supply, so very quickly after one drinks said alcohol, and blood is thin, cupula will also have a decrease in their specific gravity and will therefore float upward within their respective canal.
This is an excitation in both horizontal canals, in pairs who aren’t usually excited together, because if one horizontal canal is depolarised, this produces an opposite state of hyper-polarisation in their other. Dual excitation of both, in this example, horizontal canals, usually produces an experience of vertigo and nausea, body’s way of attempting to rid themselves of a toxin.
Just as canals detect angular acceleration, saccule and utricle detect linear acceleration. Hair cells here are embedded into rock, not cupula, this makes sense, as if we want to measure gravity, we don’t use feather, stone seems more apt, someone gravity can act on. Mass is needed.
Said rock is known as otoconial mass, made out of otoconin, a protein, with calcium carbonate, held together with a gel containing various proteins made during embryogenesis. So essentially our birth otoconial mass is with us throughout a life.
Gravity can operate on otoconial mass. In many ways inner ear crystals symbolically represent outer world, moving with them in conjunctions of opposites, motions who certain psychologists believe provide initial physical experience, gravity stage, from whom many subsequent mental introjects derive their quality.
Let’s say that I jump up and down, as I go up gravity is acting more upon me, displacing rock, otoconial mass, downwards, and so when I come back down, mass floats back up, and I have a moment of weightlessness, outside of gravity. This feeling comes from otoconial mass floating up.
For some a microscopic raft of otoconial mass can become dislodged, and they’ll float around vestibulum, eventually ending up in their lowest point, known as posterior canal. Because gravity is operating on us always, we are able to not only detect translational movement, or acceleration, but static head tilt.
If I tilt head left I’m getting a vestibular message. As I do this they effect both otolith organs, but mainly, in this sense, utricle, because head is no longer horizontal, thus displacing otoconial mass and creating a response. If I move forward, otoconial mass is displaced backward.
Otoconia have been called grains, stones, and crystals, every single crystal of inner ear is in fact multiple crystals composed of organic and inorganic compounds. Otolith crystals exist in a state of dynamic equilibrium with their fluid environment, incipiently registering effects of gravity upon brain.
Otoconial mass does not transform into a finished state who would remain unaltered throughout a life, but churn their own flux within labyrinth wherein every detail of their structure works to accommodate human audition’s existence.
Otoconia are said to mark a threshold of human identity, across whom pass processes enabling us to have our being. We can determine a fish’s age by ‘reading’ their otoconial rings.
How many people have set out on a walk in order to listen to sun?
Throughout my research into anatomical natures of human inner ear, I spent a lot of time with 19th century physicist Ernst Mach. Said research seems to have returned of their own accord.
Among many others, Mach studied semicircular canals, conducting ingenious experiments in order to deduce otolith’s abilities and roles in perceiving horizontal and vertical motion.
He also, after reading The Waste Books of polymath George Christoph Lichtenberg, in which ‘I think’ was switched out for ‘It thinks’, began to deny ego’s centrality, claiming that self equates to a series of interrelated conscious representations and sensations, continual flux.
However, what prompts this return is Mach’s recounting of a childhood memory when he heard sun hissing.
Sun’s electromagnetic nature seems quite well known, who seems less well known is semicircular canals’ relationship and sensitivity to electromagnetic fields
Seemingly we can’t listen to sun with auditory labyrinths, but might we listen with semicircular canals, wherein listening is a kind of balance?
This has led me to imagine a fourth semicircular canal, a hyper-canal, one who is primed to receive vibration in a higher dimension.
Semicircular canals are our most ‘suitable’ organ to receive electromagnetism, because endolymph is sensitive to electromagnetic fields.
We might equivocate this fourth canal as a hologrammatic projection, encircling our bodies, engaged in imagined experiences of celestial sound walks, listening to Oort clouds’ electromagnetic fields.